


背景介绍
蛋白质功能和结构分析的研究是一个非常重要的课题,因为此类研究为新药的开发或疾病的治疗提供了关键信息。X 射线晶体学通常用于高分辨率分析蛋白质结构,然而,蛋白质通常是具有复杂结构的相对不稳定的生物聚合物,结晶过程作为先决条件通常是一项具有挑战性的任务。蛋白质是具有复杂结构的不稳定生物聚合物。为了了解结晶过程,已经开发并成功建立了Derjaguin-Landau-Verwery-Overbeek(DLVO)相互作用模型,其中包括弱相互作用力。 然而,蛋白质结晶过程的许多参数,如pH值、温度和溶剂组成,只能凭经验确定。另外,已证明将纳米材料添加到蛋白质溶液中可以促进结晶过程,特别是通过加速蛋白质分子的成核,这是结晶的先决步骤。
研究出发点
碳基石墨烯纳米材料由于其优异的化学稳定性和较大的蛋白质吸附表面积,特别适合促进蛋白质结晶。石墨烯是碳的同素异形体,其中碳原子成二维 (2D) 纳米结构,并且很容易获得具有明确定义的特性,包括出色的电子、 热、化学、 和物理特性。 此外,石墨烯已经广泛用于各种应用,例如传感器、 药物输送,和光催化剂。GQDs是一种很有前途的生物功能纳米材料,因为它们具有良好的生物相容性和丰富的边缘,可以用各种化学官能团进行功能化,这些特征与其大表面积相结合,使它们成为启动或加速蛋白质结晶过程的良好候选添加剂材料。
全文速览
基于此,全北国立大学金勇珠团队报道了通过在蛋白质溶液中加入石墨烯量子点 (GQDs),可以加速溶菌酶蛋白质的结晶过程。他们的 X 射线晶体结构分析表明,GQDs和溶菌酶之间的相互作用增强了蛋白质晶体的稳定性,降低了从随机卷曲位点获得的柔韧性参数,清楚地表明GQDs是理想的纳米材料成核剂,并表明GQDs可能与蛋白质结合,并在结晶过程中起关键作用。文章以“Graphene Quantum Dots as Nucleants for Protein Crystallization”为题发表在Cryst. Growth Des.上。
图文解析
通过使用明场光学显微镜每30 min计算晶种数来研究GQDs在蛋白质结晶过程中的作用,持续 6 h。与没有GQDs的溶菌酶溶液相比,在混合GQDs (0.5 mg/mL) 的溶菌酶溶液中观察到更多晶种的形成,如图 1a中的红色箭头所示。请注意,当醋酸钠(pH 4.8)缓冲溶液与GQDs混合时,当NaCl浓度从6%降低到3%时,仍观察到溶菌酶晶种,而在相同的条件下不含GQDs的缓冲溶液未观察到晶种形成。当溶菌酶溶液中包含0.5 mg/mL GQDs时,形成的晶种数量是不包含GQDs时的约6倍(图1b),之后证实了包含GQDs不仅增加了形成的晶种总数,而且增加了结晶速率,如图1c所示,在结晶过程开始的 2 h内,含有GQDs的溶菌酶溶液中晶种的数量突然增加,而不含GQDs的溶液中晶种的数量增加可以忽略不计。2 h后,在有和没有GQDs的溶液中形成的晶种数量的增加率变得相似,两种情况都显示晶种数量随时间线性增加。这一结果表明溶液中的GQDs添加剂在结晶过程开始后 2 h内耗尽。
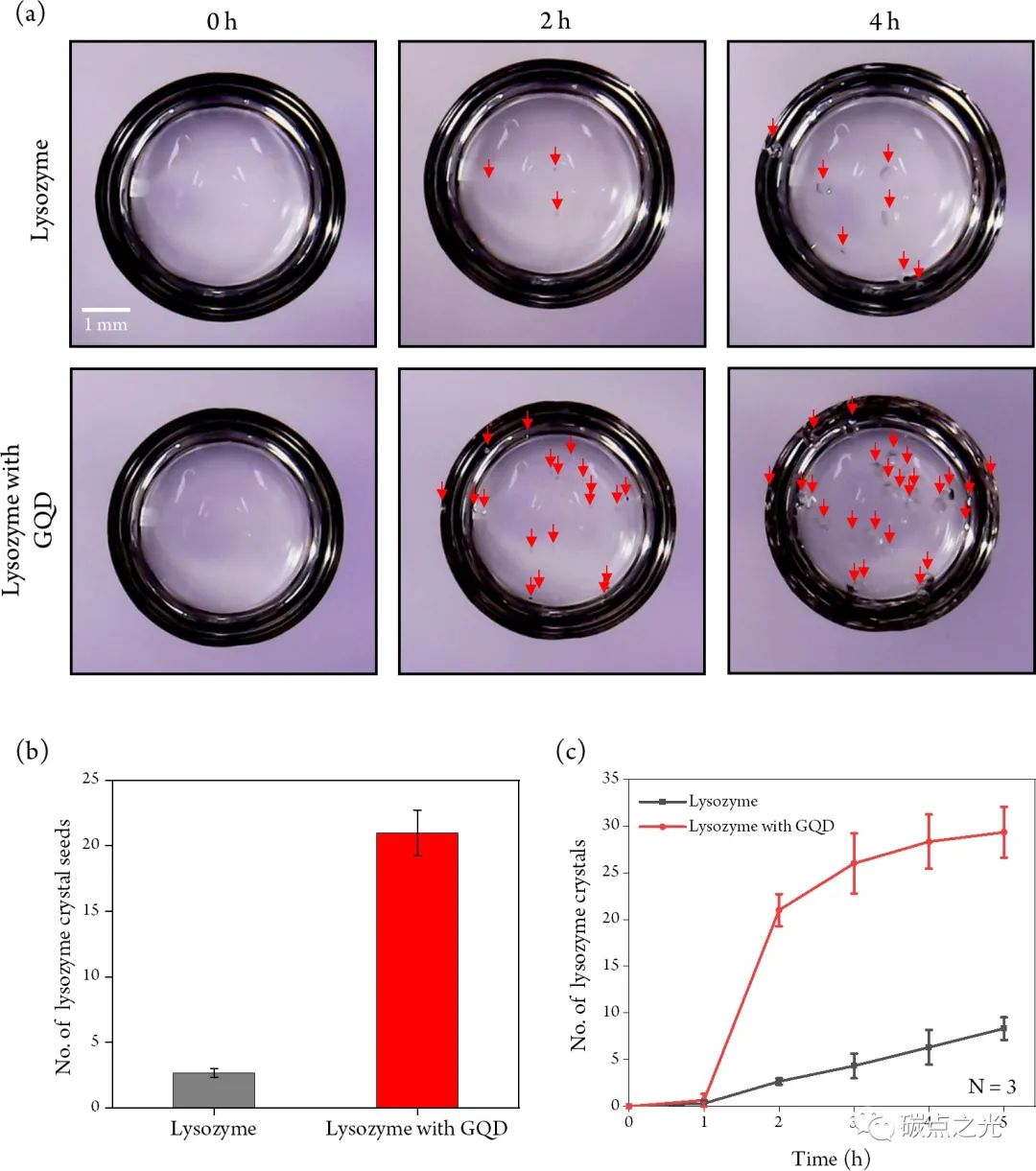
图1(a) 在不存在或存在GQDs (0.5 mg/mL) 的情况下溶菌酶结晶(红色箭头)的明场显微镜图像。(b) 孵育 2 h后晶种的数量。(c) 作为时间函数的晶种数量。
将GQDs引入溶菌酶溶液导致确定的流体动力学直径增加约30%(图2a )。随着GQDs浓度增加到超过0.8 mg/mL,体积分布曲线的高斯拟合表明流体动力学直径从不含GQDs的溶菌酶溶液的1.1 nm增加到与GQDs添加剂混合的溶菌酶溶液的1.47 nm(图 2b)。将溶菌酶的流体动力学尺寸增加归因于它与GQDs的结合,并以这种方式形成晶体成核种子。对观察结果的另一种可能解释是离子强度对DLS测量的流体动力学尺寸的影响。综合所有这些结果,特别是从溶菌酶-GQDs溶液中测量到的流体动力学尺寸随着GQDs浓度的增加而增加,表明GQDs与溶菌酶结合。
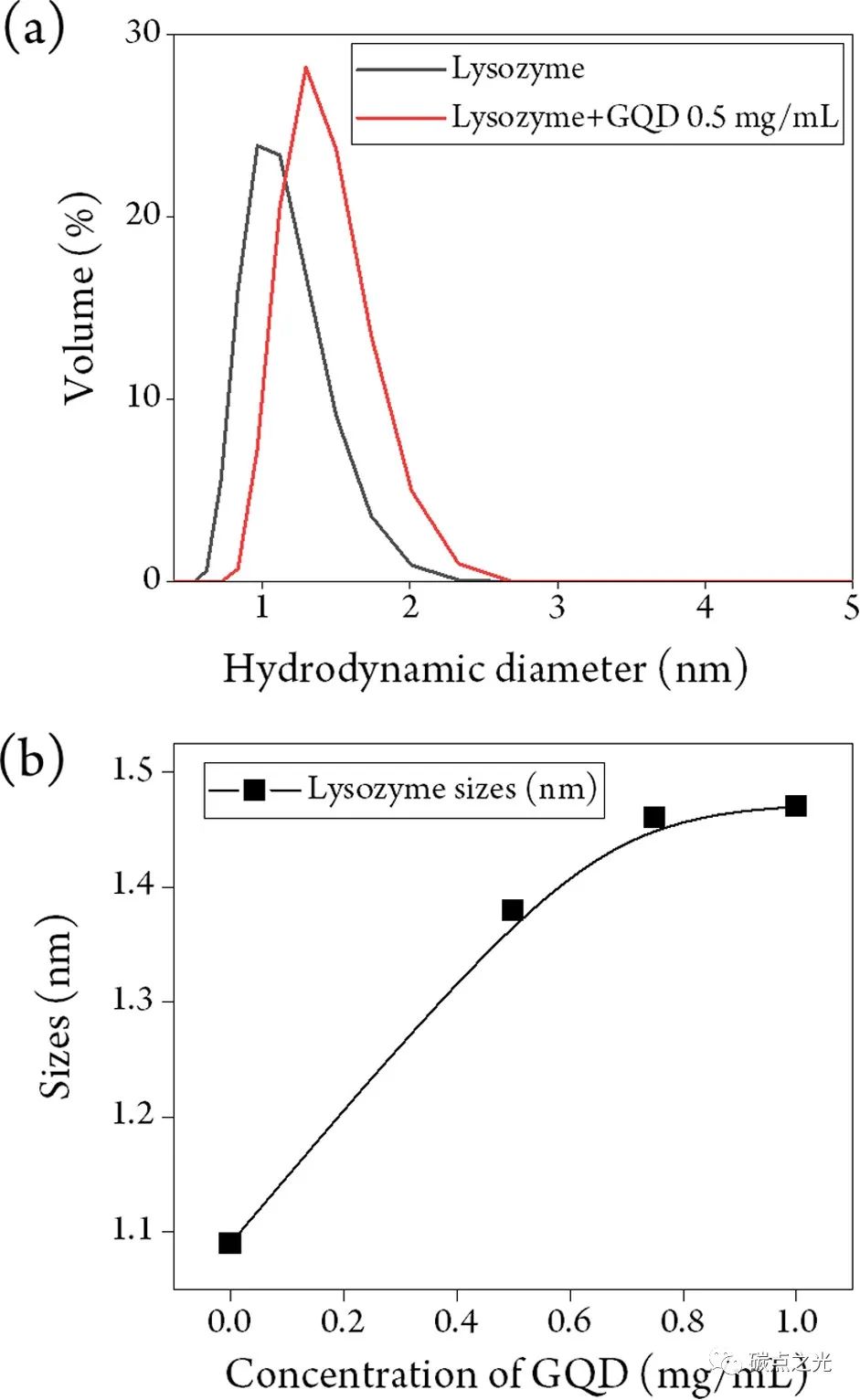
图2 在不存在或存在GQDs(0.5 mg/mL) 的情况下对溶菌酶的DLS分析显示 (a) 体积分布曲线和 (b) 作为GQDs浓度函数的流体动力学大小。
从没有GQDs生长的溶菌酶晶体中,确实没有观察到对应于GQDs的荧光信号(图 3a)。具有溶菌酶特异性峰的拉曼光谱证实,明场图像中显示的聚集体对应于溶菌酶晶体(图3e,绿线)。同时,在荧光图像中,在0.25 mg/mL GQDs存在下生长的晶体的荧光几乎不可见,这意味着大部分GQDs荧光因与溶菌酶蛋白结合淬灭。相比之下,当使用0.5和1 mg/mL 的较高GQDs浓度时,在与明场图像匹配的位置获得了显示溶菌酶晶体的清晰荧光图像,如图 3c和 d所示,从共聚焦扫描图像中,我们证实GQDs纳米材料均匀地分散在溶菌酶晶体中。GQDs对应的背景荧光信号强度与GQDs浓度不成正比,如图 3e.。例如,用1 mg/mL GQDs生长的晶体(蓝线)的背景荧光比用0.25 mg/mL GQDs生长的晶体(黑线)高约3倍。为了确认溶菌酶晶体内部GQDs的存在,将晶体切成两块,并将横截面朝向物镜放置以获得晶体横截面的荧光图像。晶体横截面的荧光图像与明场图像非常吻合,确保晶体中存在GQDs。
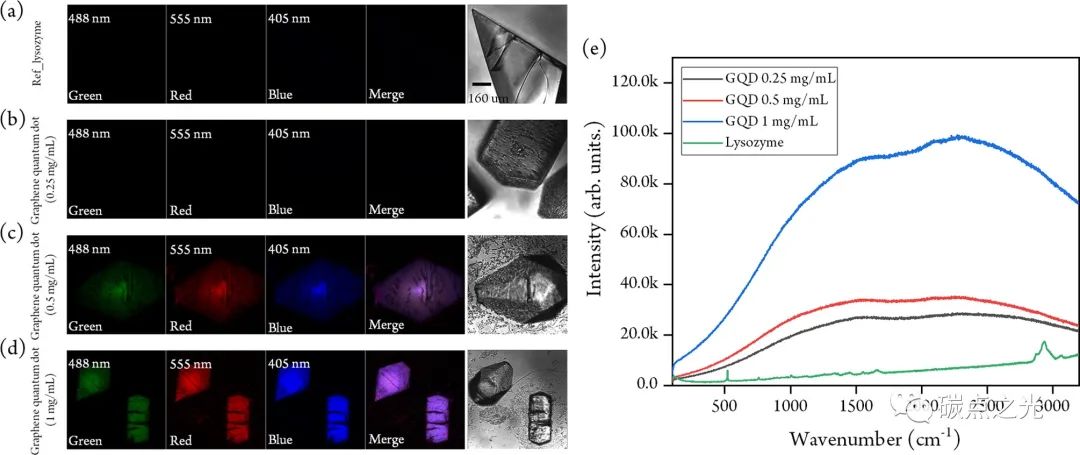
图 3. 使用 (a) 0、(b) 0.25、(c) 0.5 和 (d)1 mg/mL GQDs生长的溶菌酶晶体的共聚焦扫描显微镜图像。相应的明场图像显示在右侧。比例尺为160 μm。指示了激发波长。( e )在不存在或存在GQDs的情况下生长的溶菌酶晶体的拉曼光谱。
为了解释x射线晶体结构数据,采用了7DER模型,其结构分辨率与本工作的x射线数据相似。此外,模型中使用的结晶缓冲液在pH值(4.5)和溶液组分方面与本工作相似(图4a)。在7DER中,包含残基T47 – G49和S60- W62的两个环通常表现出很高的灵活性(图4b),这是由于它们的原子的热波动。然而加入GQDs的溶菌酶晶体结构显示了这些环的稳定性水平,如图4c所示。之后为了比较,使用PHENIX对+GQD和−GQD溶菌酶结构进行了结构分析,与+GQD结构相比,-GQD结构在T47- G49和P70 – R73残基的两个环中显示了更高的b因子值。在加入GQDs后,溶菌酶的结构降低(图4),支持GQDs的加入增加了溶菌酶蛋白的稳定性的观点。值得注意的是,与-GQDs相比,+GQD溶菌酶结构没有显著的构象变化,尽管它们的稳定性(+GQD)增加。
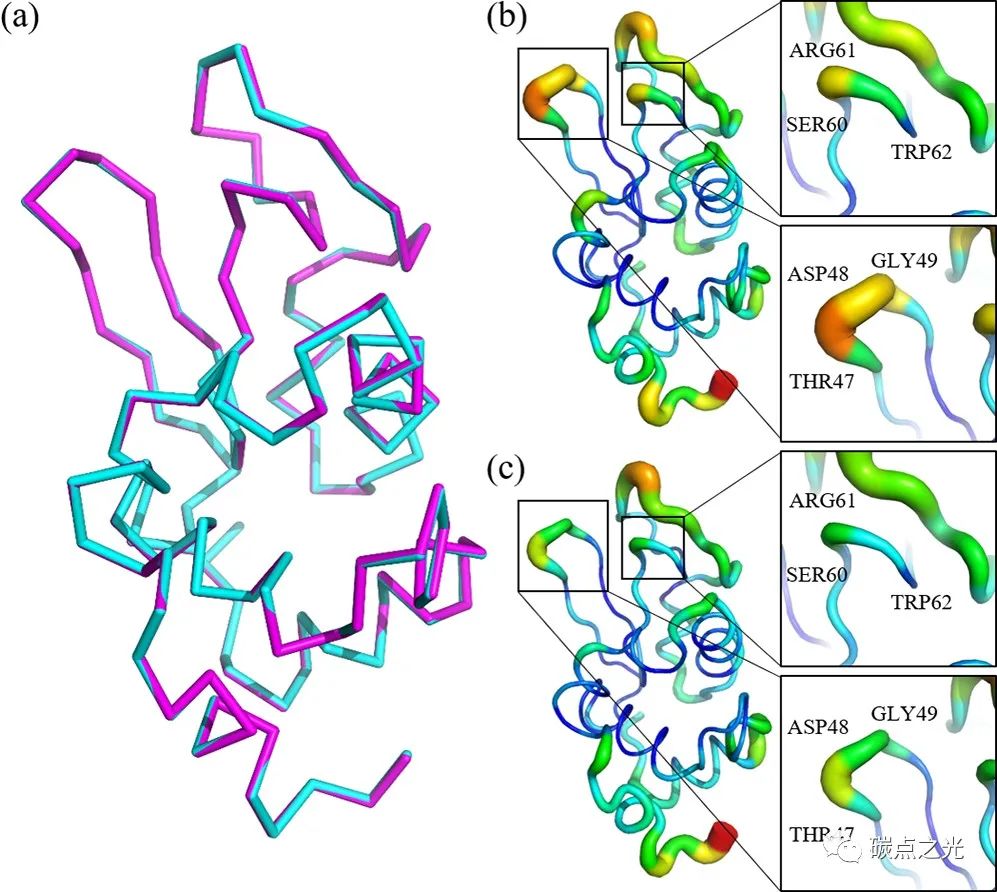
图 4. (a) 溶菌酶结构 PDB ID 的叠加:7DER和用GQDs结晶的结构,分别表示为青色和洋红色带。(b) 7DER和 (c) 溶菌酶加GQDs的 B 因子表示,在 PyMol 中生成。框表示高度灵活的区域。
总结与展望
生长适合 X 射线的蛋白质晶体是 X 射线晶体学中最困难的部分,因为它耗时且对环境条件敏感。在本研究中,研究表明包含GQDs可有效缩短蛋白质晶体成核和生长所需的时间。当包含GQDs时,DLS 测量的溶菌酶的流体动力学直径增加了约 30%,这意味着溶菌酶与GQDs相互作用的发生。在GQDs存在下生长的溶菌酶晶体的共聚焦荧光显微镜成像清楚地证实了晶体中GQDs的存在。尽管有这样的结果,X 射线晶体学分析没有显示用GQDs生长的溶菌酶晶体的结构参数与没有GQDs生长的溶菌酶晶体的结构参数之间有任何显着差异,除了当包含GQDs时蛋白质的稳定性增强。尽管在本研究中我们无法直接展示GQDs如何嵌入晶体中,但添加GQDs被认为是促进蛋白质晶体结构分析的有前途的方法。
文献链接:https://doi.org/10.1021/acs.cgd.1c00910
本文来自碳点之光,本文观点不代表石墨烯网立场,转载请联系原作者。